
Bağışıklık Sistemi ve Lenfoid Organlar
Bağışıklık sistemi bütün vücuda yayılmış, birincil işlevi organizmayı mikroorganizmaların ve yabancı maddelerin yayılmasından ve zararlı etkilerinden korumak olan çeşitli hücre ve yapılardan oluşur. Bağışıklık sisteminde bulunan hücreler bütün organizmanın kendi makromoleküllerini ve yabancı makromoleküllerden ayırt etme yeteneğine sahiptirler ve yabancı maddelerin (bağımsız moleküllerin), mikroorganizma parçalarının, hatta organizmada gelişen kanser hücrelerinin yıkımını ya da etkisizleşmesini ayarlar. Seyrek olarak İmmun sistem kendi organizmasının makromoleküllerine karşı tepkime geliştirip otoimmün hastalıkların ortaya çıkmasına neden olabilir. İmmun sistem içinde hem lenf düğümleri, dalak gibi bağımsız yapılar, hem de immun tepkimelere katılan kan, lenf ve bağ dokularında bulunan lenfositler, graniilositler ve mononükleer fagosit sistem hücreleri gibi serbest hücreler yer alır. Bağışıklık sisteminin diğer önemli bir bileşeni, sadece lenfoid organlarda değil deri gibi yabancı antijenlerle yoğun olarak karşılaşan organlarda da bulunan antijen sunucu hücrelerdir. Bağışıklık sisteminde bulunan hücreler birbirleri ile ve diğer sistemlerin hücreleriyle, öncelikle sitokinler olarak isimlendirilen sinyal proteinleri aracılığıyla haberleşirler.
Lenfoid Organlar
Bağışıklık yanıtında yer alan başlıca yapılar timus, dalak ve lenf düğümü gibi lenfoid organlardır. Esas olarak nodüler yığılmayla oluşan ve lenfoid nodül adı verilen daha küçük lenfoid doku birikimleri sindirim (tonsiller, Peyer plakları, apendiks), solunum, üriner ve üreme sistemleri mukozalarında mukoza-ilişkili lenfoid dokuyu (MALT) oluşturur. Geniş lenfoid doku dağılımı ve hücrelerin kan, lenf ve bağ dokularındaki sabit dolaşımı (resirkülasyonu), immünokompetan hücreler yardımıyla vücut için önemli bir gözetim ve savunma sistemi oluşturur. Tüm lenfositler kemik iliğinden köken alırlar; ancak T lenfositler ileri olgunlaşmalarını timusta gerçekleştirirken, B lenfositleri kemik İliğini olgun hücreler olarak terk ederler. Bu yüzden kemik iliği ve timus öncelikli veya merkezi lenfoid organlar olarak tanımlanırlar. Lenfositler bu organlardan çoğalır ve farklanmalarını tamamladıkları lenfoid organlaranodüller, tonsiller, apendiks, göç ederler.
Bağışıklık Tepkimelerinin Temel Tipleri
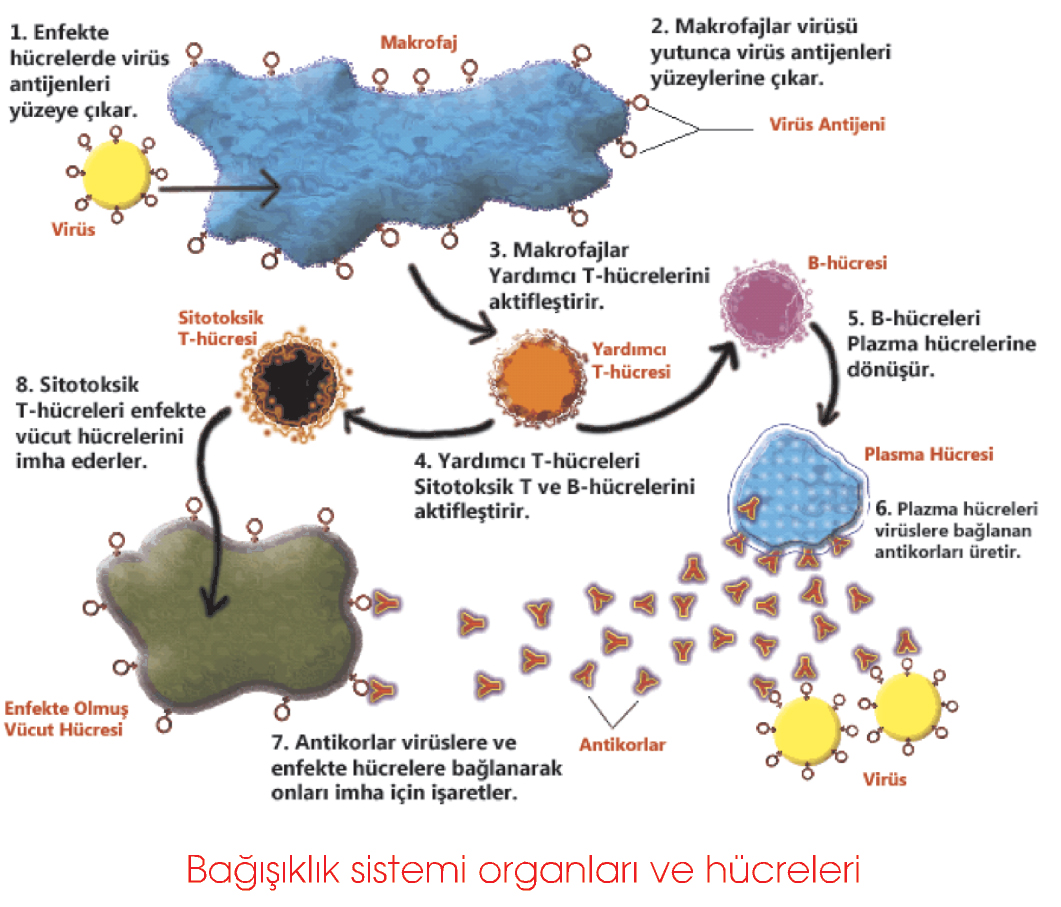
Hücresel bağışıklıkta hücreler mikroorganizmalara, yabancı (tümör ve transplantlar) ve virüsle enfekte hücrelere karşı tepkime verir ve öldürürler. Bağışıklığın bu türünde başlıca T lenfositler ya da T hücreleri etkindir. Humoral (salgısal) bağışıklık, yabancı maddeleri etkisiz hale getirip ortadan kaldıran, antikor adı verilen dolaşan glikoproteinlerin varlığıyla ilişkilidir. Antikorlar, B lenfositler veya bu hücrelerden köken alan plazma hücreleri tarafından üretilirler. Bağışıklık yanıtı aynı zamanda doğal ve edinilmiş (adaptif, kazanılmış) bağışıklık şeklinde ayrılabilir. Doğal bağışıklık; yanıtı en hızlı, özgül olmayan ve patojen ile daha önceki bir temastan bağımsız en basit tepkimedir. Doğal bağışıklık yanıtında görevli başlıca hücreler fagositler (makrofaj ve nölrofiller) ve doğal öldürücü (NK) hücrelerdir. Bağışıklık sisteminin bir parçası olmamalarına karşın, vücut içi boşlukları döşeyen örtü epiteli hücrelerinin ve derinin oluşturduğu engel, organizmayı yabancı molekül ve mikroorganizmaların girişine karşı korur. Edinilmiş bağışıklık daha ilkel olan doğal yanıtın gelişmiş özgülleşmiş biçimidir. Edinilmiş bağışıklık daha etkin ve özgül olup bir belleğe sahiptir. Aynı patojenle ikinci kez karşılaşıldığında, birinci saldırıdan daha hızlı ve etkin bir tepkime oluşturulur. Edinilmiş bağışıklık tepkimesi, yalnız doğal bağışıklığın hücrelerini değil antikor yapımını da içerdiğinden daha yavaş gelişir.
İmmünojenler ve Antijenler
Bağışıklık sisteminin karşılaştığı yabancı madde immünojen olarak davranır yani organizmada bir tepkimeye yol açar. Yanıt; hücresel, hümoral veya sıklıkla her iki tipi de içeren tarzda olabilir. İmmünojenler; bakteri, tümör hücresi gibi bütünlüğünü koruyan hücreler içinde veya protein, polisakkarid, nükleoprotein gibi makromoleküllercle bulunabilirler. Daha özgül bir tanımlamayla; bir bağışıklık yanıtı oluşturamasa bile, bir antikorla tepkime veren immünojene antijen (Yunanca; anti -karşı, + genin) denir. Çoğu immünojen aynı zamanda antijen olduğu İçin bu kitapta antijen terimi kullanılacaktır. Hümoral bağışıklık yanıtının özgüllüğü, antijenik determinant veya epitop denilen, antijenin küçük moleküler parçacıkları tarafından belirlenirken, hücresel yanıtın özgüllüğü, antijen sunucu hücrelerin zarındaki majör doku uyumu kompleksi (MHC) molekülleriyle ilişkili küçük peptidler tarafından belirlenir. Bakteri hücresi gibi pek çok epitopu olan bir antijen, geniş bir hümoral ve hücresel yanıt yelpazesi sergiler.
Antikorlar
Antikorlar veya diğer adlarıyla immünoglobulinler, yapımlarına neden olan antijenik epitop ile özgül olarak etkileşen dolaşan plazma glikoproteinleridir. Antikorlar, B lenfositlerinin çoğalıp ve farklanmasıyla oluşan plazma hücreleri tarafından salgılanırlar. Antikorun önemli bir işlevi, tanıdığı epitop ile özgül olarak birleşmesi ve bağışıklık sisteminin diğer üyelerine, bunun yok edilmesi gereken bir yabancı işgalci olduğunu bildirmesidir. Bazı antikorlar, hücreleri bir araya toplama ve çözünmüş antijenleri çöktürme yeteneğine sahiptirler. Mikroorganizmaların ve zararlı moleküllerin birikmesi, işgalciyi odaklayarak fagosite edilmelerini kolaylaştırır. İmmünoglobulin-M (IgM) veya G’ye (IgG) bağlı antijenler, bir plazma proteini grubu olan kompleman sistemini uyararak mikroorganizmaların parçalanmasına yol açarlar. Uyarılmış kompleman aynı zamanda bakteri ve diğer yayılmacıların fagositozunu da uyarır. Nötrofil ve makrofajlar antijene bağlı IgG’nin Fc bölgesini tanıyan reseptörlere sahiptirler ve bu nedenle özgül antikorlar antijen ya da yayılmacı ile kompleks oluşturarak fagositik hücrelerce tutulmasını sağlarlar. İnsanda beş çeşit immunoglobulin tanımlanmıştır: IgG, serum immünoglobulinlerinin %75’ini oluşturan, en fazla bulunan sınıftır. Diğer sınıflar için bir örnek oluşturduğundan ayrıntıyla tanımlanacaktır. IgG, disülfid bağlan ve kovalen olmayan güçlerle birbirine bağlı, eş yapıda 2 hafif ve 2 ağır zincir içerir. Ayrıldıklarında, ağır zincirlerin karboksil uçları kolayca kristalize olduklarından Fc (fragment crystallizable) olarak adlandırılır. Farklı immünoglobulinlerin Fc bölgeleri pek çok farklı hücrenin reseptörüyle tepkime verir. Dört adet amino ucu (ikisi hafif, ikisi ağır zincire ait) immiinoglobulinlerin Fab (fragment antigen-binding) parçasını oluşturur. Fab parçalarının amino asit dizilişleri değişken olduğundan bağışıklık yanıtının sıra dışı özgüllüğünden sorumludurlar. IgG, plasenta engelini aşarak fetusun dolaşım sistemine ulaşan ve yeni doğanı enfeksiyonlardan koruyan tek immünoglobulindir. IgA, kanda az miktarda bulunur. Gözyaşı, ağız (kolostrum), tükrük ve burun, bronş, barsak, prostat, vajina salgılarında bulunan ana immünoglobulindir. Salgılarda, salgısal (sekretuvar) IgA adında, 2 monomerik IgA molekülünün protein J adlı bir polipeptid zinciriyle birleşmesi ve diğer bir protein olan, salgısal ya da transport parçasıyla bütünleşmesi ile oluşan bir dimer biçiminde bulunur. Sekretuvar IgA, enzimlere dirençli olması nedeniyle mikroorganizmaların vücut sıvılarında çoğalmasını önler. IgA monomerleri ve protein J; sindirim, solunum, üriner yolların mukozalarındaki plazma hücrelerince salgılanırken, sekretuvar parça mukoza epitel hücreleri tarafından sentezlenir. IgM, kan immünoglobulinlerinin %10’unu oluşturur ve sıklıkla 900kDa ağırlığında bir beşli yapı (pentamer) şeklindedir. IgD ile birlikte B lenfositlerinin yüzeyinde bulunan başlıca immünoglobulindir. Bu iki sınıf immiinoglobulin hem hücre zarına bağlı hem de dolaşan formlarda bulunurlar. Lenfosit hücre zarına bağlı IgM ve IgD özgül antijenler için reseptör görevi görürler. Bu etkileşimin (zardaki Ig reseptörlerinin antijenle bağlanmasının) sonucu, B lenfositlerin çoğalıp, antikor salgılayan plazma hücrelerine farklanmasıdır. Dolaşımdaki IgM aynı zamanda, karaciğer tarafından sentezlenen bir grup proteinden oluşan kompleman sisteminin uyarılmasında da etkilidir. IgE, sıklıkla monomer şeklinde bulunur. Bu immünoglobulin, mast hücreleri ve bazofillerin hücre zarlarında bulunan reseptörlere yüksek ilgi gösterir. Plazma hücreleri tarafından salgılanmasının hemen ardından, IgE bu hücrelere tutunarak dolaşımdan uzaklaşır. Özgül bir IgE üretimine neden olmuş bir antijen ile yeniden karşılaşıldığında, mast hücresi veya bazofilin yüzeyinde oluşan antijen-antikor kompleksleri; bu hücrelerin histamin, heparin, lökotrienler, anaflaksinin eozinofil kemotaktik faktörü (ECF-A) gibi biyolojik aktif maddelerin üretim ve salınmasını uyarır. Bir allerjik tepkime, IgE ve onun üretimini uyaran antijenler (allerjenler) tarafından düzenlenir. IgD’nin özellik ve işlevleri tam olarak anlaşılamamıştır. Molekül ağırlığı 180 kDa olup kan plazmasındaki immünoglobulinlerin sadece %0.2’sini oluştur. IgD, IgM ile birlikte, B lenfositlerinin hücre zarlarında bulunur ve bu hücrelerin farklanmalarında görev alır.

B ve T Lenfositler
Farklanma yerleri ve hücre zarlarındaki ayırt edici reseptörlerin varlığıyla, lenfositler 2 ana sınıfa ve çeşitli alt sınıflara ayrılırlar. Bu reseptörler; B lenfositlerinde immünoglobulinler, T lenfositlerinde ise T hücre reseptörleri (TCR) adı verilen özel moleküllerdir. B lenfositler kemik iliğinden köken alarak orada olgunlaşırlar ve uyarılınca çoğalıp antikor salgılayan plazma hücrelerine farklanmak üzere yerleştikleri ikincil (sekonder) lenfoid yapılara kanla taşınırlar. B hücreleri, her biri özgül antijenler için reseptör işlevi gören 150.000 IgM molekülüyle donatılmıştır ve dolaşımdaki lenfositlerin %5-10’unu oluştururlar. Bazı uyarılmış B hücreleri, plazma hücrelerine dönüşmek yerine, aynı antijene ikinci kez maruz kalma durumunda hızlı tepkime veren bellek B hücrelerine (B memory7) farklanırlar. T hücreleri kan lenfositlerinin %65-75’ini oluştururlar. Kemik iliğinden köken alarak çoğaldıkları timusa göç ederIer ve kan yoluyla diğer lenfoid dokulara taşınırlar. T hücrelerinin; yardımcı (helper), sitotoksik (cytotoxic) ve bellek (memory) T hücreleri olmak üzere üç ana alt grubu vardır. Yardımcı ‘I’ hücreleri, B hücrelerinin plazma hücrelerine kırklanmalarını uyarırlar. Sitotoksik T hücreleri, yabancı ve virüs ile enfekte hücrelere karşı 2 yolla savaşırlar. Birincisinde, perforin adı verilen proteinler üretip, hedef hücre zarında delikler oluşturarak hedef hücrenin parçalanmasına yol açarlar. İkincisinde ise, programlanmış hücre ölümünü ya da apoptozisi başlatan bazı genleri uyararak hedef hücreyi öldürürler. Bellek T hücreleri, yeniden karşılaşılan antijenlere (patojenlere) hızlı bir yanıt vererek sitotoksik T hücrelerinin oluşumunu uyarırlar.
Klinik Bilgi
AIDS olarak bilinen immün yetmezlik sendromuna yol açan retrovirüs (HIV: human immunodeficiency virus), yardımcı T lenfositlerin ölümüne neden olur. Bu virüsle bulaşmış hastaların bağışıklık sistemi bu nedenle zayıfladığından, bu hastalar, sağlıklı insanlarda bulaş oluşturmayan mikroorganizmaların yol açtığı fırsatçı infeksiyonlara karşı duyarlı hale gelirler.
B ve T hücrelerinin lenfoid sistemde eşit olarak dağılmamışlardır ve timus dışında kalan lenfoid yapılarda özel bölgelerde yerleşmişlerdir. B ve T hücreleri, ışık ve elektron mikroskobu düzeylerinde morfolojik olarak birbirlerinden ayırt edilememelerine karşın immünositokimyasal yöntemlerle tanımlanabilirler. Alt gruplarının tespit ve tanımlanmalarını sağlayan farklı yüzey proteinleri (işaretleyicileri) taşırlar. Antijen ile uyarılma sonucunda, B ve T hücreleri, klon seçimi ve genişlemesi adı verilen bir dizi mitoz bölünme geçirerek çoğalırlar. B ve T hücrelerine ek olarak, tipik B ve T hücre işaretleyicilerini taşımayan doğal öldürücü hücre (natural killer cell, NK) adı verilen lenfositler vardır. Dolaşan kandaki lenfositlerin %10-15’ini doğal öldürücü hücreler oluştururlar. Daha önceden bir karşılaşma ve uyarılma olmadan virüs ile bulaşmış hücrelere ve kanser hücrelerine saldırdıkları için doğal öldürücü hücre adını almışlardır.
Antijen Sunucu Hücreler
Antijen sunucu hücreler, sınıf II majör doku uyumu moleküllerini (MHC-1I) taşımaları ile karakterizedirler. Sadece antijen sunucu hücrelere özel olmayan antijen işleme, T hücrelerinin uyarılabilmesi için bir ön gereksinimdir, çünkü bu hücreler organizmanın kendine ait proteinler ve diğer antijenlere karşı henüz kördür. T hücreleri sadece majör doku uyumu molekülleri ile birlikte peptidleri tanıyabilirlerken, B hücreleri doğrudan protein, peptid, lipid, polisakkaridler ve pek çok küçük molekülü tanıyarak bunlara karşı tepkime verirler. Endositoz ile alınmış yabancı proteinler endozom lizozom sisteminde sindirilir ve açığa çıkan küçük peptidler (10-30 amino asit) sınıf il majör doku uyumu kompleksi ile birleşirler. Enfekte hücrelerde yaşayan patojenlerden (virüs, bazı bakteri ve protozoalar) kaynaklanan proteinler, sınıf I majör doku uyumu kompleksi molekülleri ile birleşmek üzere endopkızma retikulumunun sisternalarına taşınacak küçük peptidlere (8-11 amino asit) kadar proteazomlarda parçalanırlar. Daha sonra, hem sınıf I hem de sınıf II kompleksler, T lenfositleri tarafından tanınacakları hücre zarına taşınırlar. CD4+ T hücreleri sınıf II molekülü- peptid kompleksi ile, CD8+ T hücreleri ise sınıf I molekülü- peptid kompleksi ile etkileşirler. CD4 (yardımcı) ve CD8 (sitotoksik), bazı T hücrelerinde bulunarak bu T hücre tiplerinin tanımlanmalarını sağlayan yüzey protein molleküleridir.
Majör Doku Uyumu Kompleksi
Bağışıklık sistemi kendine ait veya yabancı olanların ayrımını hücre yüzeylerindeki majör doku uyumu kompleksi (MHC) moleküllerinin varlığı ile gerçekleştirir. İlk olarak lökositlerde saptandıkları için insan lökosit antijenleri (HLA: human leukocyte antigen) olarak da anılırlar. Bu moleküller 2 sınıftır ; majör doku uyumu kompleksi I (MHCI) tüm hücrelerde bulunurken, majör doku uyumu kompleksi II (MHC-II) daha kısıtlı bir dağılımla sadece antijen sunucu hücrelerde bulunur. Majör doku uyumu kompleksi molekülleri, T hücreleri tarafından tanınmak üzere majör doku uyumu molekülü işlenmiş antijen kompleksini hücre zarına yerleştiren hücre içi bir sistem oluşturur. B lenfositleri serbest antijenler tarafından direkt olarak uyarılırlar. Bu lenfositler işlevlerini yerine getirmek için majör doku uyumu komplekslerinin yardımına gereksinim duymazlar. Majör doku uyumu kompleksi molekülleri, yapısı bireye özgü bazı genlerin ekspresyonu ile sentezlenir. Aynı molekülleri taşımayan bireyler arasında yapılan doku veya organ transplantlarının atılımı (rejeksiyonu) bu nedenledir.
Diğer bütünleyici zar proteinleri gibi, majör doku uyumu kompleksi I ve II proteinleri de poliribozomlarda sentezlendikten sonra granüllü endoplazma retikulumuna taşınırlar. Zincirler tamamlandıktan sonra, karboksii uçları retikulumun zarına gömülü kalacak şekilde granüllü endoplazma retikulumu sisternalarına sarkarlar. Bundan sonra, 2 sınıfın majör doku uyumu kompleksi proteinleri hücre zarına ulaşmak üzere farklı yollar izlerler. En önemli fark, majör doku uyumu kompleksi I proteinlerinin endozom-lizozom sisteminden geçmemesi, majör doku uyumu kompleksi II proteinlerinin ise bu yola girerek işlenmiş antijenlerle kompleks yapabilmeleridir. Majör doku uyumu kompleksi I proteinlerini içeren veziküllere proteazomlarda işlenmiş polipeptidler eklenebilirler ve bu veziküller işlenmiş antijenlerini sunmak üzere hücre zarına katılırlar.
ORGAN TRANSPLANTASYONU
Transplant (aktarım) edilen doku veya organ bireyin kendisinden alındığında otograft, tek yumurta ikizinden alındığında izograft, aynı türden alındığında homograft, farklı bir türden alındığında ise heterograft olarak sınıflandırılır. Organizma etkili kanlanma sağlandığı sürece otograft ve izograftlan kabul eder. Transplant edilen hücreler, alıcının hücreleri ile genetik olarak aynı oldukları ve yüzeylerinde aynı majör doku uyumu kompleksi moleküllerini taşıdıkları için bu vakalarda atılım (rejeksiyon, doku reddi) olmaz. Organizma, transplant edilmiş hücreleri kendinin olarak tanır ve hücresel veya hümoral bir tepkime geliştirmez. Homograft ve heterograftlarda ise, alıcıya yabancı olan majör doku uyumu kompleksi I molekülleri taşıyan hücreler vardır ve durumlarına uygun tanımlama ve işleme tabi tutulurlar. Transplant atılımı esas olarak, transplanta giren ve transplant hücrelerini ortadan kaldıran doğal öldürücü hücreler ve sitotoksik T lenfositlerinin aktivitelerine bağlıdır.
Sitokinler
Bağışıklık sisteminin karmaşık işlevleri, sitokinler adı verilen, düşük molekül ağırlıklı (8 ve 80kDa ağırlığında) çok sayıda peptid ve glikoprotein ile hassas bir biçimde düzenlenir. Sitokinler hem hücresel hem de hümoral mekanizmaları etkilerler. Sitokinler sadece bağışıklık sistemi hücrelerine değil, sinir sistemi ve endokrin sistem hücreleri gibi kendileri için reseptör taşıyan pek çok hücre üzerine etki ederler. Başta makrofajlar ve lökositler bağışıklık sistemi hücreleri olmak üzere endotel hücreleri ve fibroblastlar gibi diğer hücre tipleri tarafından da sentezlenebilirler. Lökositlerle etkileşen sitokinlere interlökinler denir. Bunlar lökositleri uyararak çoğalma ve farklanmalarını sağlarlar. Lenfokin terimi lenfositler tarafından üretilen sitokinleri ifade eder. Monosit ve makrofajlar tarafından üretilen sitokinler monokin olarak anılırlar. Çoğul etkileri olan pek çok sitokin tanımlanmıştır. Bu hücreler arası sinyal molekülleri, sadece lokal ve sistemik bağışıklık yanıtlarını değil; yangı, yara iyileşmesi, hematopoez gibi diğer biyolojik süreçleri de düzenlerler. Sitokinler otokrin düzenekle kendilerini üretmiş olan hücreler üzerinde, parakrin düzenekle kısa mesafedeki hücreler üzerinde ve endokrin düzenekle de uzaktaki hücreler üzerinde etki gösterirler. Kemotaksinler ya da kemokinler, lökositleri yangı bölgesine çekmek üzere uyaran sitokinlerdir. İnterferonlar, virüs ile bulaşmış herhangi bir hücre tarafından üretilen glikoprotein yapısındaki sitokinlerdir. İnterferonlar komşu makrofaj, fibroblast ve lenfositlerin reseptörleri ile tepkimeye girerek virus çoğalmasını engelleyen maddeler üretmelerini sağlarlar. Tümör nekroz faktörü denilen sitokin, çoğu kez lokal ve genel etkilere sahiptir. Bu etken; adezyon moleküllerinin, makrofajlar tarafından kemokin salgılanmasını, hedef hücrelerin apoptozisini uyarır ve ateş gibi sistemik etkilere yol açar. Kompleman Sistemi Kompleman sistemi; esas olarak karaciğer tarafından sentezlenen, her biri bir C harfi ve takip eden bir sayı ile tanımlanmış, ortalama 20 adet proteinin oluşturduğu bir havuzdur. Adı, bu sistemin, bazı bağışıklık işlevlerini tamamlayarak bütünleştirmesinden gelmektedir. İki yolla başlatılalabilen kompleman sistemi uyarılma süreci, başlangıç parçası yapısının değiştirilerek, tepkimenin diğer bileşenlere ilerleyici olarak aktarılmasıyla işleyen bir olaylar dizisi biçiminde gerçekleşir. Uyarılmış tamamlayıcı sisteminin önemli bir özelliği, makrofajlar üzerinde reseptörleri bulunan molekülleri bakterilerin yüzeyine bağlamasıdır. Bu olay, işgalci bakterilerin fagositozunu ve yok edilmelerini kolaylaştırır. Bakteriyi, makrofaja daha kolay tutunmak üzere hazırlayan bu işleme opsonizasyon denir. Kompleman sisteminin diğer bir görevi, bakteri hücre zarına hasar veren bir kompleks üretmesidir.
Bağışıklık Sistemi Hastalıkları
Bağışıklık sistemi, pek çok hücre tipi ve sinyal molekülünü İçeren, çok yaygın ve karmaşık düzeneklerle işler. Bu, tanımlanmış bağışıklık sistemi hastalıklarının çeşitliliğini açıklar. Bu hastalıklar 3 tip olarak sınıflandırılabilirler:
- Hastalık, bazı antijenlerin etkilerinin nötralize edilmeye çalışıldıkları esnada meydana gelen anormal ve şiddetli tepkimenin sonucunda ortaya çıkabilir. Bu abartılmış dayanıksızlık, değişik allerjik tepkimelere yol açar.
- Kompleman sistemi parçalarında veya makrofaj ve nötrofillerin fagositoz yeteneklerinde bozukluk, B ve T lenfositlerindeki anormallikler, antijenlere karşı olan tepkimenin baskılandığı, immün yetmezlik denilen patolojiye yol açarlar.
- Organizmada kendi antijenlerine saldıran T lenfositlerinin bulunması otoimmün hastalıklara yol açar. Bu durumda, organizmanın kendisi tarafından üretilen T lenfositleri dokuları etkiler hatta yok eder.
Damarlanma
Timusun arterleri organa kapsülden girer, bağ dokusu bölmeleri (septa) içinde dallanır ve derine ilerler. Arteriyoller parankime dalmak üzere korteks-medulla sınırı boyunca bölmeleri terk eder. Bu arteriyoller kortekse, yay şeklinde bir yol izleyen kapiller dallar verir, daha sonra medullaya ulaşarak venüllere dökülürler. Medulla, korteks-medulla sınırındaki arteriyollerin kapiller dallarıyla beslenir. Medullanın kapillerleri de, korteksten dönen kapillerlerin açıldığı venüllere dökülürler. Timus kapillerlerinin penceresiz bir endoteli ve çok kalın bir bazal laminası vardır. Bu kapillerler özellikle proteinlere karşı geçirgen değildir ve dolaşan antijenlerin, T lenfositlerinin üretildiği timus korteksine ulaşmalarını engeller. Medulla venleri bağ dokusu bölmelerine girer ve kapsülden geçerek timusu terk ederler. Medullada kan- timus engeli bulunmamaktadır. Timusun getirici (afferent) lenf damarı yoktur, lenf düğümleri gibi, lenf için bir filtre oluşturmaz. Timusta bulunan az sayıda lenf damarı; kan damarlarının duvarlarında, bağ dokusu bölmeleri ve kapsülde bulunan götürücü (efferent) lenf damarlarıdır.
LENF DÜĞÜMLERİ
Lenf düğümleri, lenf damarları boyunca vücutta dağınık olarak yerleşmiş, kapsüllü, yuvarlak ya da böbrek biçimli lenfoid doku içeren organlardır. Lenf düğümleri koltuk altında, kasıklarda, boyundaki büyük damarlar boyunca yerleşmişlerdir; göğüs boşluğunda ve karın boşluğunda, özellikle mezenlerlerde çok sayıdadırlar. Lenf düğümleri, başlıca mikroorganizmalara ve tümör hücrelerinin yayılmasına karşı organizmayı savunan ardarda dizilmiş bir seri filtre gibidirler. Doku sıvısı kökenli lenf, dolaşıma girmeden önce en az bir lenf düğümünden süzülür. Lenf düğümlerinde dışbükey bir yüzey İle arterler ve sinirlerin girdiği, venler ve lenfatiklerin organı terk ettikleri hilum (hilus) adı verilen iç bükey bir çukurluk vardır. Organı dıştan saran bağ dokusu kapsülü organın içine trabekula denilen bölmeler gönderir. Her lenf düğümünün dış korteks, iç korteks ve medülla adlı bölümleri vardır.
Korteks
Yapısal olarak dış korteksin devamı olan iç korteks çok sayıda T lenfosit ve eğer varsa az sayıda nodül içerir.
Medülla
Medülla, medülla kordonları denilen, B lenfositler ve plazma hücrelerini İçeren, kordon şeklinde, dallanmış iç korteks uzantılarından oluşur. Medülla kordonları, genişlemiş kapiller benzeri yapılar olan medüller lenf sinüsleriyle birbirlerinden ayrılmıştır. Medüller sinüsler lenfle dolu düzensiz boşluklardır; subkapsüler ve ara sinüsler gibi duvarları kısmen retiküler hücreler ve makrofajlarla döşelidir. Retiküler hücreler ve lifler gevşek bir ağ örgüsüyle sinüs lümeninde köprüler oluştururlar.
Klinik Bilgi
B ve T lenfositlerin işlevleri en belirgin olarak, B hücreleri, T hücreleri ya da her ikisinin birden hatalı olmasından kaynaklanan immün yetmezlik hastalıklarında ortaya çıkar.

Histofizyoloji
Lenf, dolaşım sistemine geri dönmeden önce lenf düğümlerinden geçerken yabancı partiküllerden temizlenir. Lenf düğümleri tüm organizmaya dağılmış olduğundan dokularda oluşan lenf, dolaşıma katılmadan önce en az bir lenf düğümünün içinden geçer.
Lenfositlerin Geridolaşımı (Resirkülasyon) Bir İletişim Sistemi
Vücutta oluşan tüm lenf sonunda kan dolaşımına geri döndüğünden, efferent lenfatiklerle lenf düğümünü terk eden lenfositler de sonuçta kan dolaşımına ulaşırlar. Lenfositler postkapiller ya da yüksek endotelli venül adı verilen özel kan damarlarıyla kandan ayrılarak lenf düğümüne geri dönerler. Bu venüller alışılmışın dışında, lenfositlerin aralarından geçtiği, yüksek boylu kübik hücrelerden oluşmuş bir endotel yapısı gösterirler. Yaşama süresi uzun olan bazı lenfositler bu şekilde defalarca dolanırlar. Yüksek endotelli venüller ayrıca appendiks, tonsiller ve Peyer plaklan gibi diğer lenfoid organlarda da bulunur ancak dalakta bulunmaz. Resirkülasyon diğer lenfoid organlarda bir oranda gerçekleşmekle birlikte lenf düğümlerinde en belirgindir. Lenfositlerin özel olarak yüksek endotelli venüller aracılığıyla lenfoid organlara girmeleri, lenfositlerle yüksek boylu endotel hücrelerinin yüzey moleküllerinin birbirlerini tamamlayıcı özellikte olmalarıyla gerçekleşir. Lenfositlerin resirkülasyonuyla, uydu lenf düğümlerinde lokal olarak uyarılmış lenfositler (örneğin enfekte bir parmakta), diğer lenfoid organları da olaydan haberdar edip, organizmayı infeksiyona karşı genel bir immün yanıt oluşturmaya hazırlarlar. Lenfositlerin sürekli resirkülasyonu immün sistemin vücuttaki tüm yabancı antijenlerin varlığını izlemesini olanaklı kılar. Lenfositler lenfoid organlardan geçişleri sırasında, bulaşmış bölgeden göç ederek gelmiş olan antijen sunucu hücrelerin zarlarında yerleşmiş tüm antijenlerle karşılaşırlar.
DALAK
Dalak vücuttaki en büyük lenfoid doku topluluğudur. Dolaşımdaki kanla yakın temasta bulunan çok sayıda fagositik hücre içermesi nedeniyle dalak, dolaşıma girmiş tüm mikroorganizmalara karşı önemli bir savunma bölgesidir. Dalak aynı zamanda yaşlanmış eritrositlerin dolaşımdan uzaklaştırıldığı bölgedir. Diğer lenfoid organlar gibi, dalak da kana verilen uyarılmış lenfositlerin bir üretim bölgesidir. Dalak kanda taşınan antijenlere karşı hızla reaksiyon geliştiren önemli bir immünolojik filtre ve antikor üretici organdır.
Genel Yapı
Dalak, sıkı bağ dokusu yapısında, parenkimi ya da dalak pulpasını birbiriyle devamlılık gösteren bölümlere ayıran trabekülaların uzandığı bir kapsülle sarılıdır. Dalağın orta çizgi hizasındaki hilumda kapsül, dalak pulpasına sinirleri ve arterleri taşıyan çok sayıda trabekula oluşturur. Parankimada oluşan venler ve trabekülaların bağ dokusu içinden başlayan lenfatik damarlar, hilumdan organı terkederler. Dalak pulpasında lenf damarı bulunmaz. İnsanda kapsül ve trabekülaların bağ dokusu içinde sadece bir kaç düz kas hücresi bulunur. Dalak diğer tüm lenfoid dokular gibi retiküler dokudan oluşan bir ağ içine yerleşmiş lenfoid hücreler, makrofajlar ve antijen sunan hücrelerden oluşmuştur.
Kan Dolaşımı
Hilumdan organa giren splenik arter, dallanarak bağ dokusu trabekülalarında seyreden değişik boyutlardaki trabeküler arterleri oluşturur. Trabeküler arterler bağ dokusunu terk edip parankimaya girince damarın çevresi, T lenfositlerden oluşmuş periarteriyel lenfatik kılıf (PALS) adlı bir kılıfla sarılır. Bu damarlar, santral arterler ya da beyaz pulpa arterleri olarak bilinir. Lenfositik kılıf (beyaz pulpa) damar çevresindeki seyri boyunca yer yer lenf nodülleri içeren genişlemeler gösterir ve santral arter adı verilmiş olmasına karşın lenf nodülleri içinde bu damarlar çevresel bir yerleşim gösterir. Santral arter beyaz pulpa içindeki seyri boyunca, çevredeki lenfoid dokuyu besleyen ışınsal düzenlenmiş yan dallar verir. Beyaz pulpayı terkeden santral arter dallanarak dış çapı yaklaşık 24 nm olan düz penisiller arteriyolleri oluşturur. Bazı penisiller arteriyoller sonlanmalarına yakın bölümde, ıeliküler hücreler, lenfoid hücreler ve makrofajlardan oluşan bir kılıfla sarılmışlardır. Kılıfı geçtikten sonra penisiller arteriyoller kanı sinüzoidlere (kırmızı pulpa sinüslerine) taşıyan basit arteriyel kapilerler olarak devam ederler. Bu sinüzoicller kırmızı pulpa kordonları arasında bulunan alanı doldurur. Kırmızı pulpanın arteriyel kapillerlerinden sinüzoidlerin içine doğru olan kan akımının ne şekilde gerçekleştiği henüz tam olarak açıklığa kavuşmamıştır. Bazı araştırmacılar kapillerlerin doğrudan sinüzoidlere açıldığını iddia ederlerken, diğerleri kanın önce kırmızı pulpa kordonlarındaki hücreler arasındaki boşluklardan süzülüp sinüzoidlerde toplandığını ileri sürmektedir. Birinci görüş kapalı dolaşım olarak bilinir; yani kan daima damarların içinde seyreder. İkinci görüş açık dolaşımı savunur. Bu görüşe göre dolaşım kırmızı pulpa parankiması (Billroth kordonları) içine açılır ve kan sinüzoidlere ulaşmak için hücreler arasındaki boşluktan geçer. Bugünkü bilgiler insan dalağında dolaşımın açık türden olduğunu destekler yöndedir. Kan sinüzoidlerden kırmızı pulpa venlerine doğru ilerler. Kırmızı pulpa venleri birbirleriyle birleşerek parankimayı terkedip trabekülaların bağ dokusu içine girerler ve trabeküler venleri oluştururlar. Trabeküler venlerin birleşmesiyle oluşan splenik ven hilumdan organı terk eder. Trabeküler venlerin duvarında kendilerine ait bir kas tabakası yoktur, duvarları trabeküler dokudan oluşmuştur. Bunlar trabeküler bağ dokusu içinde yerleşmiş endotelle döşeli kanallar olarak düşünülebilirler.
Beyaz Pulpa
Beyaz pulpa santral arterleri ve kılıflara ilişik lenf nodüllerini saran lenfoid dokudur. Santral arterleri saran lenfoid hücreler başlıca T lenfositlerdir ve bunlar periarteriyel lenfatik kılıfları (PALS) oluştururlar Lenf nodülleri ise başlıca B lenfositleri içerir. Beyaz ve kırmızı pulpa arasında, sinüslerin bol bulunduğu gevşek bir lenfoid doku uzanır. Bu bölge marjinal zon olarak bilinir. Burada bol miktarda aktif makrofaj ve az sayı¬ da lenfosit bulunur. Kandaki antijenlerin yoğunlaştığı bu bölge dalağın immünolojik aktivitesinde önemli bir role sahiptir. Periarteriyel lenfatik kılıfın artere yakın iç bölgelerindeki lenfositler timus bağımlı (T) lenfositlerdir. Buna karşılık marjinal zon ve nodüllerde yani periferik beyaz pulpada ise B lenfositler yerleşmiştir
Kırmızı Pulpa
Kırmızı pulpa dalak kordonlarını ve venöz sinüzoidleri içerir. Dalak kordonları retiküler liferle (tip-III kollajen) desteklenmiş gevşek bir retiküler hücre ağından oluşmuştur. Kordonlar bunlara ek olarak makrofajları, lenfositleri, plazma hücrelerini ve çok sayıda kan hücresini (eritrosit, trombosit ve granitlosiler) içerir. Dalakta sinüzoidler, uzun eksenleri sinüzoidin uzun eksenine paralel yerleşmiş yassı endotel hücreleriyle döşelidir. Bu hücreler dıştan tıpkı bir fıçının kasnakları gibi enine düzenlenmiş retiküler liflerle sarılmıştır. Enine ve uzunlamasına lifler birleşerek ağsı bir yapı oluşturarak komşu endotel hücreleri arasındaki boşlukları dolduran makrofajları ve sinüzoid hücrelerini çevreleyip sararlar. Sinüzodin çevresinde kesintili bir bazal lamina vardır. Sinüzoidleri döşeyen hücreler arasındaki aralıklar 2-3 nm çapında ya da daha dar olduğundan ancak esnek hücresel elemanlar, kırmızı pulpadan sinüzoid lümenine geçebilir. Daha önce değinildiği gibi sekonder lenfoid organlarda (yapılarda) T ya da B lenfositlerin daha yoğun olduğu bölgeler vardır.
Histofizyoloji
Dalağın en iyi bilinen işlevleri lenfosit üretimi, eritrositlerin yıkımı, kan dolaşımına giren yayılmacılara karşı organizmanın korunması ve kanın depolanmasıdır.
LENFOSİT ÜRETİMİ
Dalağın beyaz pulpası içinde üretilen lenfositler kırmızı pulpaya göç edip sinüzoidlerin lümenine ulaşarak dolaşıma geçerler.
Klinik Bilgi
Lösemi gibi bazı patolojik durumlarda, dalak miyeloid metaplaziye (kemik iliği dışındaki bölgelerde miyeloid dokuların görülmesi) uğrayarak fetal dönemdeki gibi eritrosit ve granülositleri üretmeye yeniden başlayabilir.
ERİTROSİTLERİN YIKIMI
Yaklaşık 120 günlük bir yaşam süresi olan eritrositler bunun sonunda başlıca dalakta olmak üzere ortadan kaldırılırlar. Esnekliklerinin azalması ve zarlarındaki değişiklikler ortadan kaldırılmaları için bir sinyal oluşturuyor görünmektedir. Yozlaşmakta olan eritrositler kemik iliğinde de ortadan kaldırılırlar. Dalaktaki makrofajlar ekstraselüler aralıkta parçalanan eritrositleri fagosite ederek ortadan kaldırırlar. Hemoglobinin parçalanmasıyla ortaya çıkan protein (globin) aminoasitlere kadar hidrolize edilerek yeni protein yapımında kullanılır. Hemoglobinin hem bölümünden açığa çıkan demir, transferin ile birleşip kan yoluyla kemik iliğine taşınarak, eritrosit yapımında tekrar kullanılır. Hem’İn demir ayrıldıktan sonra kalan bölümü ise bilirubine dönüştürülüp, karaciğer hücrelerince safraya atılır. Dalağın çıkarılmasından (splenektomi) sonra, kanda şekil bozuklukları gösteren, anormal eritrositlerin sayısı artar. Aynı zamanda, dalağın yaşlanmış trombositleri (plateletleri) dolaşımdan uzaklaştırdığı savına destek olur biçimde periferik kanda trombosit sayıları da artar.
ORGANİZMANIN SAVUNULMASI
Dalak, T ve B lenfositleri, antijen sunucu hücreleri ve makrofajları içerdiğinden organizmanın savunmasında önemli bir görev üstlenir. Tıpkı lenf düğümlerinin lenfi süzmesi gibi, dalak da kanı süzen bir filtre olarak işlev görür. Kana karışmış bakteri ve virüs gibi canlı ya da yapısı bozulmayan cansız partiküllerin fagosite edilmesinde, dalak makrofajları organizmada bulunan tüm fagositik hücreler arasında en aktif olanlarıdır.
MUKOZAYA EŞLİK EDEN LENFOİD DOKU
Sindirim, solunum ve genitoıiriner yollar dış ortama açık olduklarından mikrop yayılmasının en sık görüldüğü bölgelerdir. Vücudu korumak için bu yolların mukoza ve submukozasında tonsil ve ince bağısaklarda ki Peyer plakları gibi belirgin yapılan da oluşturan lenf nodülleri (lenfoid kümeler) ve yaygın lenfoid doku birikimleri izlenir. Deri de immun sistemin pek çok hücresini (lenfositler, makrofajlar, Langerhans hücreleri) barındırır. Deri ve mukozalardaki lenfoid dokular, çevreden gelen patojenler için kilit yerleşimde etkin bir savunma sistemi oluştururlar.
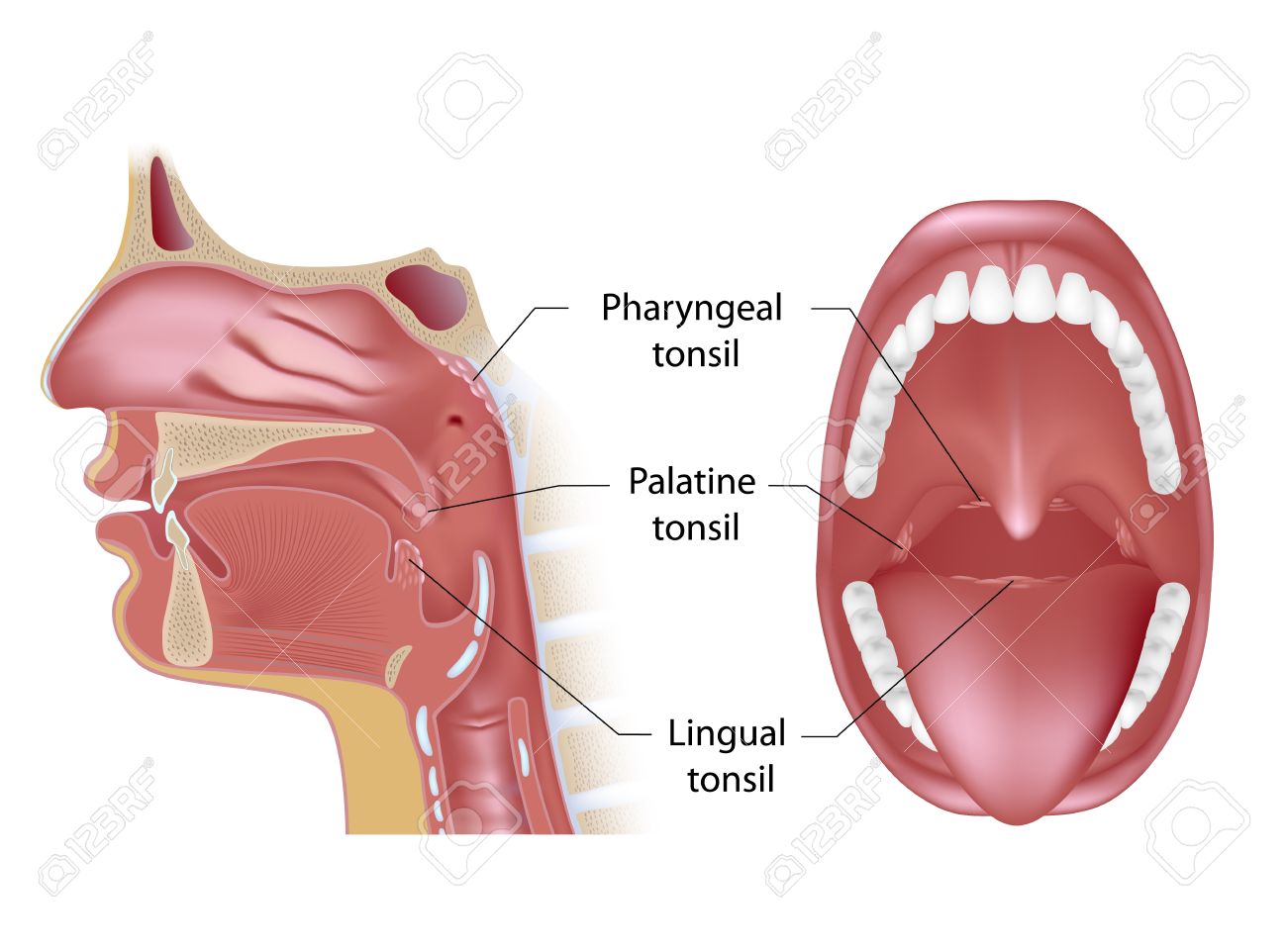
TONSİLLER (BADEMCİKLER)
Tonsiller sindirim kanalının başlangıç bölgesinde epitel altında ancak epitelle ilişkili olacak şekilde yerleşmiş, kesintili bir kapsülle çevrelenmiş lenfoid organlardır. Yerleşim yerlerine göre ağız ve larinks bölgesindeki tonsiller palatin, faringeal ve lingual tonsiller olarak adlandırılır. Tonsillerde çoğu epitel içine girmiş olan lenfositler üretilir.
Palatin Tonsiller (Damak Bademcikleri)
Farinksin ağız bölümünün yan duvarlarına yerleşmiş sağlı sollu bir çift palatin tonsil bulunur. Bu tonsillerdeki yoğun lenfoid doku, çok katlı yassı epitelin altında, genellikle germinal merkezler içeren lenf nodüllerinden oluşan bir bant şeklinde izlenir. Her bir tonsilde sayıları 10-20 arasında değişen, lümeninde dökülmüş epitel hücreleri, canlı ya da ölü lenfositler ve bakterilerin bulunduğu kripta adı verilen derin epitel girintileri vardır. Bu bölgeler tonsillitte ufak iltihabi noktalar şeklinde izlenir. Lenfoid dokuyu altındaki diğer yapılardan ayıran kapsül adı verilen bir sıkı bağ dokusu tabakası vardır. Kapsül tonsil bulaşıcılarının çevre dokulara yayılmasını önleyen bir engel oluşturur.
Faringeal Tonsil (Yutak Bademcikleri)
Farinksin arka üst bölümüne yerieşmiş tek bir tonsildir. Organ solunum yollarının tipik yalancı çok katlı silyalı prizmatik epiteliyle örtülü olup yer yer çok katlı yassı epitel alanları da içerebilir. Mukoza pililerinden oluşan bir yapıya sahip faringeal tonsilde diffüz lenfoid doku ve nodüller bulunur. Kriptası yoktur ve kapsülü palatin tonsilden daha incedir. Faringeal tonislin kronik iltihap nedeniyle hipertrofısi adenoid olarak adlandırılır.
Dil Bademcikleri
Tonsiller diğerlerinden küçüktür ve çok fazla bulunmaktadır. Dil kökünde yerleşmişlerdir ve çok katlı, yassı epitelle örtülüdür.